Abstrac
A pesar de la disponibilidad de múltiples agentes terapéuticos, la búsqueda de un nuevo manejo del dolor del dolor neuropático sigue siendo un desafío. El estrés oxidativo y la señalización inflamatoria están prominentemente involucrados en la manifestación clínica del dolor neuropático. Toxicodendron pubescens , popularmente conocido como Rhus Tox (RT), se recomienda en medicamentos alternativos como un remedio antiinflamatorio y analgésico. Anteriormente, informamos sobre las actividades antiinflamatorias, antiartríticas e inmunomoduladoras de Rhus Tox . A continuación, evaluamos la eficacia antinociceptiva de Rhus Tox en el dolor neuropático y delimitamos su mecanismo subyacente. Inicialmente, in-vitroSe realizó un ensayo con células de glioblastoma U-87 inducidas por LPS mediadas por LPS para estudiar el efecto de Rhus Tox sobre las especies reactivas de oxígeno (ROS), el estado de antioxidantes y el perfil de citoquinas. Rhus Toxredujo el estrés oxidativo y la liberación de citoquinas con la restauración de los sistemas antioxidantes. El tratamiento crónico con diluciones de Rhus Tox durante 14 días mejoró el dolor neuropático revelado como una inhibición de la alodinia fría, cálida y mecánica, junto con una mejor velocidad de conducción del nervio motor (MNVC) en el nervio constricto. Rhus Toxdisminuyó el estrés oxidativo y nitrosativo al reducir el contenido de malondialdehído (MDA) y óxido nítrico (NO), respectivamente, junto con el aumento de glutatión regulado (GSH), superóxido dismutasa (SOD) y la actividad de la catalasa en el nervio ciático de ratas. En particular, el tratamiento con Rhus Tox causó reducciones significativas en los niveles de factor de necrosis tumoral (TNF-α), interleucina-6 (IL-6) e interleucina-1β (IL-1β) en comparación con el grupo control CCI. El efecto protector de Rhus Tox contra la lesión del nervio ciático inducida por CCI en el estudio histopatológico se demostró mediante el mantenimiento de la arquitectura normal del nervio y la inhibición de los cambios inflamatorios. En general, el efecto neuroprotector de Rhus Tox en el dolor neuropático inducido por CCI sugiere la participación de mecanismos antioxidantes y antiinflamatorios.
Introducción
El dolor neuropático representa una condición clínica importante y hay una búsqueda continua de nuevos tratamientos para tratar la neuropatía 1 . Se manifiesta como anomalías sensoriales como disestesia (sensación desagradable), hiperalgesia (aumento de la sensibilidad a los estímulos nocivos), alodinia (aumento de la sensibilidad a los estímulos no nocivos) y desarrollo espontáneo del dolor 2 , 3 . El dolor neuropático tiene una patología compleja con distintos mecanismos. Los episodios globales de dolor neuropático se están intensificando debido al aumento del envejecimiento de la población, el aumento de la supervivencia de la quimioterapia contra el cáncer y el aumento progresivo de la incidencia de diabetes mellitus 1. A pesar de la aparición de nuevas tecnologías de descubrimiento de fármacos y los avances en el campo de la neurociencia, el problema del manejo del dolor neuropático con remedios seguros y efectivos aún no se ha resuelto 4 , 5 . Muchos ensayos controlados aleatorios han presentado la eficacia clínica de los opioides, gabapentinoides, antidepresivos tricíclicos, inhibidores de la recaptación de serotonina y ligandos de los canales de calcio en el tratamiento del dolor neurogénico. Sin embargo, estos agentes no son suficientes para controlar el dolor neuropático. La eficacia de los fármacos antineuropáticos disponibles es limitada debido a la aparición de varios efectos secundarios y al alivio inadecuado o retardado del dolor 4 , 6 . La necesidad de descubrir nuevas modalidades de tratamiento para las condiciones neuropáticas aún prevalece.
Varios factores contribuyen a la sensibilización periférica y al inicio del dolor neuropático. El aumento del estrés oxidativo, el aumento de la permeabilidad vascular y la liberación de diferentes mediadores inflamatorios, incluida la sustancia P y el péptido relacionado con el gen de la calcitonina producido por terminales nociceptivos, la formación y / o liberación de bradicinina, prostaglandinas, factores de crecimiento y citoquinas conduce a la aparición de dolor neuropático 7 . Recientemente, los agentes antiinflamatorios y antioxidantes como la N-acetil carnitina y el ácido alfa lipoico se están investigando como medicamentos complementarios para el tratamiento del dolor neuropático 8 .
Los productos naturales o las medicinas complementarias y alternativas se utilizan con frecuencia para controlar los trastornos neurológicos crónicos como el dolor neuropático 4 . Pocos estudios preclínicos sobre remedios homeopáticos también han demostrado la eficacia de los remedios homeopáticos en el tratamiento del dolor neuropático 9 , 10 . Las encuestas basadas en la evidencia han revelado que los pacientes con dolor crónico prefieren usar tratamientos a base de hierbas para estas condiciones dolorosas 11 , 12 . Varios estudios preclínicos y clínicos han informado sobre el efecto beneficioso de las plantas medicinales en la terapia de la neuropatía dolorosa 4 , 8 , 13 , 14 , 15 ,16 , 17 . Toxicodendron pubescens P. Mill, perteneciente a la familia anacardiaceae, se conoce como Rhus toxicodendron o Rhus tox (RT) en medicinas alternativas 18 , 19. La RT es un remedio homeopático de uso común para el tratamiento de afecciones inflamatorias, dolor reumático, fiebre tipo tifoidea y afecciones de la membrana mucosa 20 . Estudios experimentales han demostrado que la RT posee inmunomoduladores 21 , antiinflamatorios 19, 22 , antiartríticos 23 , 24 y actividad contra el melanoma 25.. Recientemente, un estudio observacional prospectivo en pacientes con cáncer de mama ha demostrado que la RT puede disminuir el dolor y la rigidez en las articulaciones en mujeres con cáncer de mama temprano 26 .
La lesión quirúrgica de los nervios periféricos en animales experimentales se realiza generalmente para producir modelos animales que se asemejan al dolor neuropático humano 27 . En este sentido, el modelo de lesión por constricción crónica (ICC) parece estar bien establecido y se utiliza con frecuencia modelo animal para el estudio del dolor neuropático 27 , 28 . En este modelo, la ligadura unilateral suelta del nervio ciático imita el estado de dolor neuropático patológico en los seres humanos 29 . En este informe, demostramos los efectos antinociceptivos de la RT en el dolor neuropático utilizando ensayos invitro e in vivo . Inicialmente, in-vitrose realizó un estudio con la línea celular de glioblastoma primario U-87 para delinear el efecto de la RT en la producción de ROS inducida por LPS, el estado de antioxidantes y los niveles de citoquinas. Además, se evaluó la eficacia in vivo de la RT en el dolor neuropático utilizando un modelo animal bien caracterizado de dolor neuropático inducido por lesión por constricción del nervio ciático en ratas. Diversos paradigmas de pruebas de comportamiento (frío, calor y alodinia mecánica), parámetros fisiológicos (MNCV), estimaciones bioquímicas (MDA, NO, GSH, SOD, catalasa, TNF-α, IL-1β, IL-6) e histopatología se llevaron a cabo para explorar Los mecanismos analgésicos subyacentes de la RT en el dolor neuropático experimental. La gabapentina sirvió como control positivo en este estudio.
Resultados
Efecto de la RT en el estrés oxidativo inducido por LPS en células de glioblastoma U-87
El tratamiento de células U-87 con LPS dio como resultado un aumento notable en la viabilidad celular. Considerando que, el tratamiento de células con RT a concentraciones probadas disminuyó la viabilidad celular inducida por ROS mediada por LPS de una manera dependiente de la dosis ( Fig. 1A ). No hubo una citotoxicidad considerable debido al tratamiento de RT en ausencia de estimulación con LPS (datos no mostrados). Para anular la interferencia de la citotoxicidad de la RT, probamos las concentraciones de RT que carecían de cualquier efecto prominente en la viabilidad celular. La estimulación de células con LPS ha dado como resultado niveles reducidos de SOD y actividad de catalasa en células de glioblastoma primario U-87. Pero después de la adición de RT (1 × 10 −8 ; 1 × 10 −12 ; 1 × 10 −24 y 1 × 10 −30) en LPS, las células tratadas previamente ofrecieron un aumento significativo en SOD (*** P <0 .001="" actividad="" c="" catalasa="" comparaci="" con="" control="" de="" en="" font="" las="" lps.="" lulas="" n="" nbsp="" p="" tratadas="" y="">La RT mostró un efecto dependiente de la dosis en estos sistemas antioxidantes ( Fig. 1B, C ). El nivel de producción de ROS en células U-87 después de la estimulación con LPS se evaluó mediante análisis de citometría de flujo mediante tinción con DCFH-DA. Tras la estimulación con LPS, aproximadamente el 70% de las células U-87 se revelaron como ROS positivo. La disminución significativa (*** p <0 10="" c="" con="" concentraci="" de="" del="" dependiente="" despu="" en="" font="" la="" lps="" lulas="" n="" nbsp="" observ="" positivas="" pretratadas="" ros="" rt="" s="" se="" tratamiento="" y="">−8 ; 1 × 10 −12 ; 1 × 10 −24 y 1 × 10 −30). El porcentaje de células ROS positivas se redujo en RT (1 × 10 −8 ; 1 × 10 −12 ; 1 × 10 −24 y 1 × 10 −30 ) células tratadas, respectivamente. El H 2 O 2 se utilizó como control positivo para la generación de ROS ( Fig. 1D, K ).
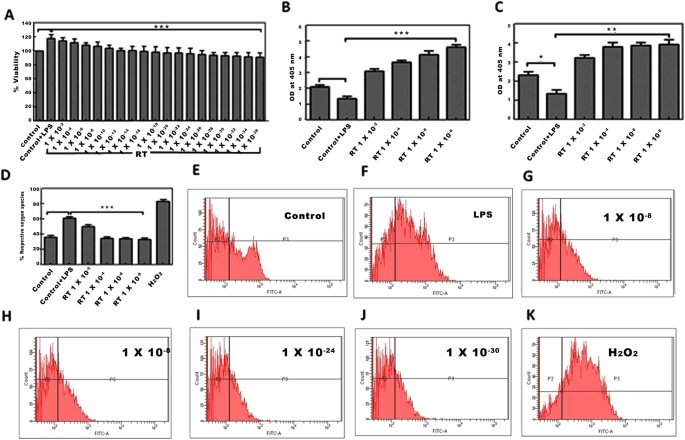
Efecto de la RT en la actividad de ROS, SOD y catalasa mediada por LPS en células U-87. ( A ) Ensayo de viabilidad celular de MTT después de la exposición a RT en células U-87 pretratadas con LPS. *** p <0 font="" nbsp="">( B ) Detección bioquímica de SOD y ( C ) catalasa después del tratamiento con RT en células U-87 pretratadas con LPS. ( D ) Las células pre-tratadas con LPS (500 ng / ml durante 20 min) se trataron con diversas concentraciones de RT antes de la estimación de ROS. La representación gráfica de la población de ROS positiva analizada mediante tinción con DCFH-DA utilizando un citómetro de flujo (E – K)fue la representación gráfica del recuento de células en Control, Control + LPS, RT (1 × 10 −8 , 1 × 10 −12 , 1 × 10 −24y 1 × 10 -30 ) y H 2 O 2 (control positivo) tratados U-87 células, respectivamente. Los datos representan la media ± SEM de 3 experimentos independientes. ** p <0 font="" nbsp="" y="">p <0 c="" comparaci="" con="" en="" font="" las="" lps.="" lulas="" n="" tratadas="">
Efecto de la RT en la expresión de citoquinas proinflamatorias inducida por LPS en células de glioblastoma U-87
Para comprobar el efecto de la RT en las citocinas proinflamatorias, se midió el nivel de TNF-α, IL-1β, IL-6 e IL-10 en células U-87 pretratadas con LPS. El tratamiento de las células con LPS (500 ng / ml durante 20 min) dio como resultado un aumento significativo en el nivel de varias citoquinas en comparación con las células no tratadas. En comparación con el control de LPS, el tratamiento de células pretratadas con LPS con RT (1 × 10 −8 ; 1 × 10 −12 ; 1 × 10 −24 y 1 × 10 −30 ) resultó en dosis dependiente y significativa disminución de los niveles de TNF-α (* P <0 .05="" e="" font="" il-1="" il-6="" nbsp="" p="">Curiosamente, el tratamiento de RT casi bajó el nivel de citoquinas al nivel basal ( Fig. 2A – D ).
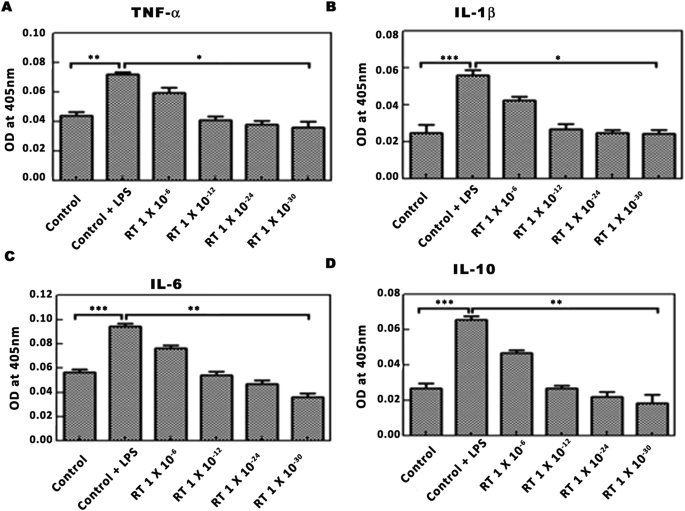
Efecto de la RT en las citocinas proinflamatorias estimuladas con LPS en células U-87. Las células pretratadas con LPS (500 ng / ml durante 20 min) se trataron con diversas concentraciones de RT antes de la estimación de las citoquinas. ( A - D ) Son la representación gráfica de la densidad óptica de los sobrenadantes celulares a 405 nm para TNF-α, IL-1β, IL-6 e IL-10. Los datos representan la media ± SEM de 3 experimentos independientes. ** p <0 font="" nbsp="" y="">p <0 c="" comparaci="" con="" en="" font="" las="" lps.="" lulas="" n="" tratadas="">
Efecto de la RT en la alodinia fría, cálida y mecánica en el dolor neuropático inducido por CCI
Efecto de la RT sobre la alodinia fría, tibia y mecánica en ratas durante el dolor neuropático inducido por CCI se representa como la Fig. 3A-C . La constricción del nervio ciático en los animales causó un desarrollo significativo de alodinia fría (Fig. 3A ), alodinia cálida (Fig. 3B ) y alodinia mecánica (Fig. 3C ) en comparación con el grupo de operación simulada, evaluado mediante una prueba de agua fría y tibia. Prueba de agua y prueba de cabello de Von Frey, respectivamente.
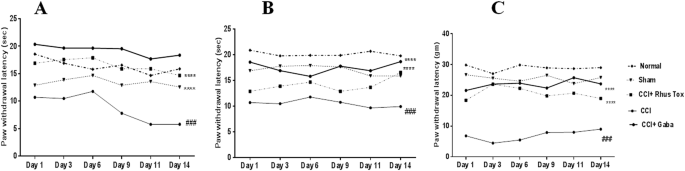
La RT mejoró los estímulos fríos, cálidos y mecánicos inducidos por la alodinia en el dolor neuropático inducido por CCI en ratas. Los datos se expresaron como la media ± SEM (n = 8). La significación estadística se determinó mediante medidas repetidas ANOVA (análisis de varianza) seguido de la prueba post hoc de Bonferroni, ### p <0 comparaci="" con="" de="" el="" en="" font="" forma="" grupo="" n="" nbsp="" operado="" simulada.="">Mientras que, la diferencia estadísticamente significativa en los grupos administrados con RT y gabapentina en comparación con el grupo control con ICC se representó como *** p <0 font="" nbsp="">La p <0 .05="" considerada="" estad="" font="" fue="" significativa.="" sticamente="">
La latencia de retirada de la pata (PWL) con respecto a la alodinia fría disminuyó significativamente en las ratas con control CCI en comparación con las ratas operadas de forma simulada (8,7 ± 1,1 seg. Vs. 16 ± 0,4 seg, P <0 font="" nbsp="">La RT ( dilución 1 × 10 −12 ) elevó significativamente el PWL en comparación con los animales control con CCI (13 ± 0,3 seg. Vs. 8,7 ± 1,1 seg; F (5, 20) = 5.641, P <0 .001="" font="" nbsp="">Estos efectos fueron similares y menos potentes que la gabapentina (60 mg / kg / día, po .). La gabapentina aumentó el PWL en respuesta a estímulos fríos hasta 19 ± 0,4 segundos en comparación con el grupo de control CCI 16 ± 0,5 (P <0 fig.="" font="" nbsp="">3A . El efecto de la RT en la alodinia cálida después de la cirugía con ICC se representa como la Fig. 3B. El CCI indujo una disminución significativa en el PWL (11 ± 0.3 segundos) en comparación con el grupo operado simulado y el grupo normal. El tratamiento con RT dio lugar a una elevación significativa de PWL de hasta 14 ± 0,5 segundos en comparación con el grupo de control CCI (P <0 como="" en="" fig.="" font="" la="" muestra="" nbsp="" se="">3B .
Se determinó el umbral de retirada de la pata (PWT) utilizando pelos de Von Frey para evaluar la alodinia mecánica en ratas. El PWT en ratas de control CCI (6.9 ± 0.7 g) fue significativamente menor en comparación con las ratas del grupo simulado. Los tratamientos con RT y gabapentina aumentaron significativamente el PWT en comparación con las ratas con control CCI como se muestra en la Fig. 3C .
Efecto de la RT sobre el MNCV en el dolor neuropático inducido por CCI en ratas
El MNCV se estimó en el 14 ° día. CCI indujo una reducción significativa en el MNCV. En comparación con el grupo operado simulado que tiene MNCV igual a 23.2 ± 3.8 mm / s, las ratas inducidas por CCI tuvieron MNCV de 15.9 ± 2.1 mm / s (P <0 .001="" font="" nbsp="">Los tratamientos con RT y gabapentina aumentaron el MNCV hasta 23.9 ± 2.07 y 25.3 ± 5.92 mm / seg respectivamente. Sin embargo, estos efectos de la RT y la gabapentina no fueron estadísticamente significativos (Fig. 4 ).
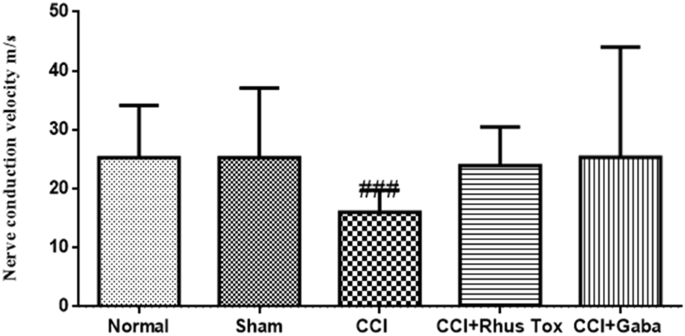
RT mejoró el MNCV en el dolor neuropático inducido por CCI en ratas. Los datos se expresaron como la media ± SEM (n = 8). La significación estadística se determinó por un método ANOVA (análisis de varianza) seguido de la prueba post hoc de Bonferroni. ### p <0 cci.="" comparaci="" con="" el="" en="" font="" grupo="" inducido="" n="" por="">
Efecto de la RT sobre el estrés oxidativo en el dolor neuropático inducido por CCI
La extensión de la peroxidación lipídica en el homogeneizado de nervio medido como MDA fue significativamente mayor en el nervio ciático ligado en comparación con el grupo operado simulado (91 ± 18 Vs. 50 ± 6,6 μg / mg de proteína; P <0 font="" nbsp="">De manera similar, el nivel de óxido nítrico se elevó significativamente en el grupo de control de CCI en comparación con el grupo simulado (P <0 font="" nbsp="">El estrés oxidativo inducido por CCI causó niveles significativamente reducidos de GSH e inhibió la SOD y las actividades de catalasa en los homogeneizados de nervios en comparación con las ratas operadas de forma simulada. La administración oral de RT durante 14 días atenuó significativamente el aumento inducido por CCI en los niveles de MDA y NO. El tratamiento con RT también restableció los niveles de actividad GSH y SOD. El efecto restaurador de la RT en la actividad de la catalasa no fue estadísticamente significativo. La gabapentina mejoró notablemente el estrés oxidativo en ratas inducidas por CCI. Además, los efectos de la RT en la reducción de los niveles de MDA y NO fueron comparables con los efectos del fármaco estándar gabapentina( Tabla 1 ).
Efecto de la RT en las citoquinas proinflamatorias en el dolor neuropático inducido por CCI
El CCI indujo un aumento notable en los niveles de citoquinas proinflamatorias. Los niveles tisulares de TNF-α, IL-6 e IL-1β en el grupo inducido por CCI fueron 56 ± 2.6, 44 ± 4.8 y 64 ± 6.8 pg / mg respectivamente. Considerando que, en el grupo de operación simulada, los niveles de TNF-α, IL-6 e IL-1β fueron 26 ± 0.4, 22 ± 0.3 y 22 ± 0.3 pg / mg. El grupo inducido por CCI tenía niveles significativamente más altos de citoquinas proinflamatorias en el tejido nervioso (P <0 font="" nbsp="">El tratamiento de RT durante 14 días disminuyó notablemente los niveles de TNF-α a 25 ± 1.1 pg / mg, IL-6 a 20 ± 1.4 pg / mg y para IL-1β hasta 20 ± 1.4 pg / mg de proteína (p <0 .001="" font="" nbsp="">Estos efectos de la RT en las citoquinas proinflamatorias fueron similares a los efectos ejercidos por la gabapentina (Fig. 5 ). La RT ejerció un efecto inhibidor más prominente en los niveles de IL-1β en comparación con el efecto de la gabapentina.
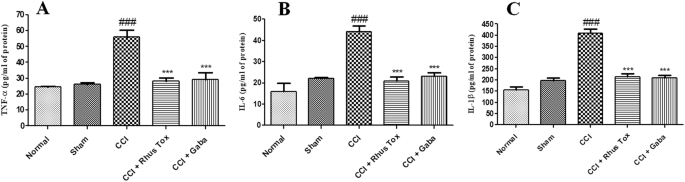
La RT redujo la liberación de citoquinas proinflamatorias en el dolor neuropático inducido por CCI. Los datos se representan como media ± SEM (n = 8). La significación estadística se analizó mediante un análisis de varianza de una vía (ANOVA) seguido de la prueba post hoc de Bonferroni. p <0 .05="" considerado="" estad="" font="" fue="" nbsp="" significativo.="" sticamente="">La diferencia estadística del grupo de control de CCI en comparación con el grupo operado de forma simulada se representó como ### p <0 .001.="" font="" nbsp="">Considerando que, la diferencia estadística de RT o grupo tratado con gabapentina en comparación con el grupo control CCI se representó como, *** p <0 font="">
Efecto de la RT en las alteraciones histológicas inducidas por CCI en el nervio ciático
El CCI indujo cambios histológicos característicos en el nervio ciático. Las alteraciones, como la inflamación de la fibra nerviosa, la infiltración de células inflamatorias, el desajuste de la fibra y la activación de células neurogliales como las células satélite y las células de Schwann sugieren el daño del nervio ciático. El tratamiento de animales con RT o gabapentina protegió el nervio de los cambios inducidos por el CCI. Las muestras de nervios de ratas que recibieron estos tratamientos revelaron que tanto la RT como la gabapentina habían reducido los desajustes estructurales, junto con la inhibición de la fibra nerviosa y la inflamación de las células neurogliales ( Fig. 6 y Tabla 2 ).
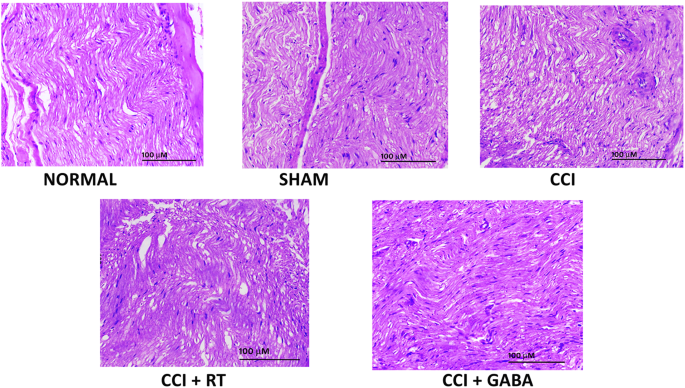
RT inhibió las alteraciones histológicas inducidas por CCI en las secciones longitudinales del nervio ciático en ratas.
Las secciones longitudinales del nervio ciático de los grupos normal, simulado, control con CCI, CCI + RT (1 × 10 12 ; 0,1 ml / día, po) y CCI + gabapentina (60 mg / kg, po) se muestran como Fig. . 6 , respectivamente. El estudio histopatológico de cortes de ratas normales demostró la arquitectura normal del nervio ciático sin perturbaciones inflamatorias. Las secciones obtenidas de las ratas operadas de forma simulada también revelaron una estructura normal sin cambios marcados. Sin embargo, las ratas con control de CCI mostraron inflamación de la fibra nerviosa, desajuste de la arquitectura de la fibra, infiltración de células inflamatorias junto con alteraciones en las células satélite y células de Schwann (Tabla 2 ). Tratamiento con RT (1 × 10 −12; 0,1 ml / día, po) o gabapentina (60 mg / kg / día, po) mostraron un efecto de mejora contra los cambios inducidos por el CCI en el nervio ciático de ratas, como lo demuestra la reducción de las alteraciones inflamatorias y el trastorno estructural (aumento 100 ×, escala barra, 1000 µm).
Discusión
El dolor neuropático inducido por CCI en ratas representa comportamientos dolorosos característicos como la hiperalgesia y la alodinia, lo que valida su idoneidad para la evaluación de medicamentos antineuropáticos 29 , 30 , 31 . En este modelo, el dolor neuropático es causado por una lesión primaria y una disfunción del sistema nervioso somatosensorial 3 , 14 . Se ha demostrado sistemáticamente que las ultra diluciones de RT poseen actividades analgésicas, antiinflamatorias 22 e inmunomoduladoras 21 en los modelos experimentales. Investigamos los efectos de la RT utilizando in vitro.modelo de estrés oxidativo inducido por LPS y liberación de citoquinas del cultivo de células de glioblastoma U-87 para determinar la ultra dilución exacta que inhibe el estrés oxidativo inducido. La ultra dilución que se encontró que era la más efectiva para reducir el estrés oxidativo en las células nerviosas se probó para determinar la eficacia antineuropática contra el dolor inducido por CCI.
La eficacia de la RT para inhibir el estrés oxidativo inducido por LPS en las células U-87 fue evidente en nuestro estudio. RT 1 × 10 −12 y 1 × 10 −15diluciones redujeron el porcentaje de células ROS positivas y también disminuyeron la expresión de citoquinas proinflamatorias (TNF-α, IL-1β, IL-6 e IL-10) junto con una disminución En los niveles de SOD y catalasa. Estos hallazgos significan el potencial in vitro de las ultra diluciones de RT para proteger las células neuronales del estrés oxidativo inducido. Dos Santos et al . 19 ha informado que la actividad antiinflamatoria de la RT es máxima cuando su dilución homeopática corresponde a 1 × 10 −12.La dilución se probó contra el edema de pata inducido por carragenina en ratas. Se encontró un patrón similar de eficacia máxima en nuestros estudios anteriores sobre RT, donde evaluamos su efecto antiinflamatorio, inmunomodulador y antiartrítico 21 , 22 , 23 , 24 . Los presentes hallazgos in vitro que indican la efectividad de la RT para reducir el estrés oxidativo y la liberación de citoquinas proinflamatorias concuerdan con estos informes.
A lo largo de los 14 días posteriores a la cirugía con ICC, las ratas mostraron síntomas de comportamiento característicos del dolor neuropático similares a los síntomas clínicos. El umbral de retirada de la pata (PWT) y la latencia de retirada de la pata (PWL) contra los estímulos mecánicos y térmicos se utilizaron para la determinación del dolor neuropático. El tratamiento de RT efectivamente elevó estos umbrales de dolor, lo que indica la reducción de las sensaciones de dolor neuropático. En línea con los datos publicados que indican las actividades antiinflamatorias 22 y analgésicas 26 , nuestro estudio demuestra que la RT también reduce eficazmente el dolor neuropático.
El CCI induce desequilibrio hipóxico isquémico en los metabolitos secundarios de las fibras nerviosas afectadas y produce estrés oxidativo 32 , 33 . La degeneración de la fibra nerviosa y la disminución de la energía nerviosa reducen el MNCV. La RT mejoró la reducción mediada por el daño nervioso en el MNCV después de la ICC en el nervio ciático, lo que indica su efecto neuroprotector. El estrés oxidativo y la inflamación son eventos interconectados que conducen a lesiones nerviosas y dolor persistente 34 , 35 . Se ha informado que diversos extractos de plantas, sus fracciones aisladas o fitoconstituyentes ejercen su actividad antineuropática a través de los mecanismos antiinflamatorios y antioxidantes 35 , 36 , 37 ,38 , 39 , 40 . Las concentraciones de MDA y NO se miden comúnmente para estudiar la extensión de las especies reactivas de oxígeno 34 . Las ROS como superóxido, NO y peroxinitrito tienen un papel importante en el desarrollo de respuestas neuroinflamatorias 41 . También se ha informado un aumento en la peroxidación lipídica después de la ICC del nervio ciático 41 . La eficacia de los captadores de radicales libres para la reducción de la peroxidación lipídica sugiere la participación de las ROS en la sensibilización nerviosa y la producción de alodinia 42 .
NO es la molécula de señalización vital que tiene un papel importante en el dolor central y periférico 43 . La participación del óxido nítrico endógeno en el desarrollo del dolor neuropático inducido por CCI está bien documentada en estudios anteriores 44 . El aumento significativo en los niveles de NO en el nervio ciático sugirió la regulación al alza de NO y su papel en la producción y mantenimiento del dolor neuropático 44 ,45. En el presente estudio, se observó un aumento significativo en los niveles de MDA y NO en el nervio con constricción crónica. Denota daño elevado a las macromoléculas, como las proteínas y los lípidos, que pueden causar dolor neuropático. La administración crónica de RT resultó en la disminución de los niveles de MDA y NO, lo que indica la actividad de peroxidación anti-lipídica junto con la inhibición del daño tisular nitrosante y el dolor neuropático sucesivo.
Nuestros hallazgos están en línea con estudios previos que informaron la disminución en el contenido de GSH del nervio ciático después de la ICC en ratas 46 . La catalasa y la SOD son las principales enzimas antioxidantes que ayudan a eliminar los radicales libres y ofrecen protección contra el estrés oxidativo 46 , 47 . En nuestro estudio, la disminución en el nivel de catalasa y SOD después de la ICC en las ratas es indicativo de aumento del estrés oxidativo o disminución de los mecanismos de defensa antioxidante. Estos resultados están en sintonía con informes anteriores y refuerzan la contribución del estrés oxidativo en el desarrollo del dolor neuropático 47 , 48 , 49. La disminución mediada por el estrés oxidativo en los niveles de GSH, SOD y catalasa se restauró en las ratas CCI mediante el tratamiento crónico con RT-dililución (1 × 10-12 ). Por lo tanto, la disminución mediada por RT en el estrés oxidativo inducido por CCI podría ser uno de los mecanismos implicados en su actividad antinociceptiva en el dolor neuropático.
Diferentes citoquinas, incluyendo TNF-α IL-1β e IL-6 se liberan después de la lesión del nervio y contribuyen al desarrollo de dolor neuropático 34 . El daño del nervio periférico induce la liberación de TNF-α rápidamente de los macrófagos, las células de Schwann y los mastocitos 26 , 50 . TNF-α se asocia con una disminución del umbral del dolor y se informa que los tratamientos anti-TNF-α alivian el dolor inducido por CCI en ratas 51. En este estudio, observamos una disminución significativa en el nivel de TNF-α en el nervio ciático en el grupo tratado con RT en comparación con el grupo control CCI. Este hallazgo reveló que la RT moduló la neuroinflamación en el nervio periférico a través de su acción anti-TNF-α y podría ser uno de sus mecanismos anti-nociceptivos. Se sabe que la IL-1β se expresa en neuronas nociceptivas y tiene un papel importante en varios modelos de dolor 52 , 53 . La IL-1β aumenta la sensibilización al dolor neuropático a través de su acción en las neuronas adyacentes 34. Observamos un aumento significativo en la producción de IL-1β en el nervio ciático después de la ICC en ratas. El tratamiento de animales con RT inhibió la producción de IL-1β, lo que puede explicar su efecto antinociceptivo. La IL-6 es una citoquina proinflamatoria que tiene un papel importante en el desarrollo de dolor inflamatorio y neuropático después de una lesión del nervio periférico. Los resultados del presente estudio revelaron que la RT tiene capacidad para disminuir el nivel de IL-6, lo que significa el efecto anti-IL-6 de la RT en el modelo de dolor neuropático inducido por CCI. Los hallazgos generales a través de la estimación bioquímica indican que la RT alteró favorablemente el perfil de citoquinas en el modelo experimental de dolor neuropático.
En resumen, este estudio demostró el potencial antinociceptivo de las diluciones de RT en el modelo validado de dolor neuropático periférico inducido por CCI. Curiosamente, se encontró que la RT era eficaz para reducir no solo la nocicepción térmica sino también la alodinia mecánica en ratas. La RT demostró notables efectos antioxidantes y antiinflamatorios en el nervio ciático y mostró una actividad potencial de eliminación de radicales libres que reduciría su actividad antineuropática. El estrés oxidativo atenuado y las vías inflamatorias pueden contribuir al potencial terapéutico de la RT en el tratamiento del dolor neuropático. Aunque, los resultados del presente estudio sugirieron el efecto antineuropático de la RT, se justifican más estudios preclínicos y clínicos para confirmar estos efectos. Varios otros mecanismos bioquímicos pueden estar involucrados en el efecto anti-neuropático mediado por RT. Los resultados del presente estudio sugieren el efecto anti-nociceptivo de la RT contra el dolor neuropático y merecen una validación adicional de su eficacia en diversas afecciones dolorosas.
Métodos
Productos quimicos
El polvo seco autenticado de hojas de Rhus Tox (Familia: Anacardiaceae) se obtuvo del laboratorio de farmacopea homeopática, Ghaziabad, Uttar Pradesh, India. La gabapentina fue dotada por los laboratorios Mylan, India. Kits de Cytokine ELISA Ready SET-Go para TNF-α (Cat: 837324-22: Lote No. E09479-1645), IL-1β (Cat: 887013-22; Lote No. E09323-1645) e IL-6 (Cat: 837064-22: El lote No. E09358-1645) se adquirió de e-Biosciences Incorporation, EE. UU. El lipopolisacárido (LPS) de Escherichia coli O55: B5 (Cat: L2880; Lote No. 025M4040V) se compró a Sigma-Aldrich, EE. UU.
Preparación de extracto etanólico RT y diluciones.
Se siguió el procedimiento prescrito en la monografía de la farmacopea homeopática de la India para la preparación del extracto de RT y sus ultra-diluciones, excepto las sucesiones características utilizadas en la preparación de las diluciones homeopáticas. Se pulverizó polvo seco y grueso de hojas de RT. El polvo exactamente pesado (10 g) se mezcló con 100 ml de etanol (70%) y se mantuvo en la jarra de vidrio para maceración en frío hasta 7 días con agitación ocasional durante cada día 36 , 54 . El 8 º día, mezcla se filtró a través de papel de filtro Whatman y el filtrado se utiliza como un extracto alcohólico de RT. Se prepararon varias diluciones de extracto en etanol para obtener las concentraciones finales de RT de 1 × 10 −2 , 1 × 10 −4 , 1 × 10 −6., 1 × 10 −8 , 1 × 10 −10 , 1 × 10 −12 , 1 × 10 −14 , 1 × 10 −16 , 1 × 10 −18 , 1 × 10 −20 , 1 × 10 −22 , 1 × 10 −24 , 1 × 10 −26 , 1 × 10 −28 , 1 × 10 −30 , 1 × 10 −32 , 1 × 10 −34 , 1 × 10 −36 (Fig. 7 ).
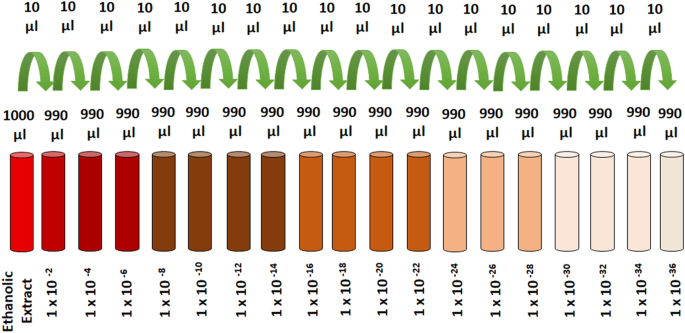
Esquemas para la preparación de diluciones de RT.
Ensayo de viabilidad celular MTT
El estudio de citotoxicidad de RT en células U-87 inducidas por ROS mediado por LPS se realizó utilizando el ensayo de viabilidad MTT [3- (4,5-dimetiltiazol-2il -) - bromuro de difenil tetrazolio] como se describió anteriormente 5 , 35 . Aproximadamente, 1 x 10 4 células se sembraron por triplicado en placas de cultivo de tejidos de 96 pocillos y se dejó que alcanzaran una confluencia del 80%. Las células U-87 se trataron con 500 ng / ml de LPS durante 20 minutos para inducir ROS. Los medios que contenían LPS se reemplazaron con medios nuevos y las células se trataron con diferentes concentraciones de RT (1 × 10 −2 –1 × 10 −36).) por más de 24 h. Luego, se agregaron soluciones de MTT (0.05 μg / μl) diluidas en PBS a cada pocillo. Las placas se incubaron durante la noche a 37 ° C para permitir la formación de cristales de formazán púrpura. Posteriormente, se añadió una solución de detergente a cada pocillo para solubilizar los cristales y se incubó durante 30 minutos a 37ºC. La intensidad del color formado después de disolver los cristales de formazán en DMSO se midió espectrofotométricamente utilizando un lector de microplacas (Berthold Technologies, Alemania) a 570 nm. Cada punto de datos se realizó por triplicado y todos los ensayos se realizaron tres veces. Los datos se representaron como el porcentaje (%) de viabilidad frente al control.
Cultivo celular y tratamiento.
La línea celular de glioblastoma humano U-87 se mantuvo en medio águila modificada de Dulbecco (DMEM) suplementado con suero bovino fetal al 10% (FBS), 100 U / ml de penicilina, 100 μg / ml de estreptomicina, 1,5 mM de L-glutamina en el Atmósfera humidificada de 5% de CO 2 a 37 ° C. Las células U-87 se cultivaron en matraces de cultivo (75 cm 2 ) y el medio se cambió cada día alternativo. Después de una confluencia del 80%, los medios se reemplazaron con medios nuevos que contenían LPS (500 ng / ml) durante 20 minutos para inducir la producción de especies reactivas de oxígeno (ROS). Se agregaron varias concentraciones de RT en células pre-tratadas con LPS durante otras 24 h antes de realizar los siguientes experimentos. La concentración de etanol en el medio de ensayo final fue inferior al 0,1%. El peróxido de hidrógeno (H 2 O 2) a la concentración fija (10 µM) se trató durante 30 minutos para producir ROS y se usó como control positivo.
Determinación de ROS, SOD, actividad de catalasa y citoquinas en células U-87
Las células U-87 pretratadas con LPS (500 ng / ml durante 20 min) se trataron con diferentes concentraciones de RT antes de la estimación de ROS. El análisis por citometría de flujo de la producción de ROS por 2'-7'-diclorodihidrofluoresceína diacetato (DCFH-DA) se realizó con un citómetro de flujo (FACS Canto II, Becton & Dickinson, CA, EE. UU.) Como Eruslanov y Kusmartsev 37 describieron anteriormente con algunas modificaciones. . El H 2 O 2 se utilizó como control positivo para la generación de ROS. La concentración de SOD y el nivel de catalasa en células U-87 pretratadas con LPS se midieron utilizando los métodos reportados anteriormente 38 , 39. La cuantificación de citoquinas, incluyendo TNF-α, IL-1β, IL-6, IL-10 en células U-87 pretratadas con LPS, se realizó en sobrenadantes de cultivo celular utilizando 50 μg de proteína mediante los kits ELISA obtenidos comercialmente según las instrucciones del fabricante. 5 , 38 .
Los animales
Se utilizaron ratas Wistar albinas adultas de ambos sexos (170–220 g) para el presente estudio. Los animales se obtuvieron de las instalaciones de la casa de animales del Instituto RC Patel de Educación e Investigación Farmacéutica, Shirpur. Los animales se mantuvieron en jaulas de polipropileno ventiladas bajo las condiciones estándar (25 ± 2 ° C, ciclo de luz / oscuridad de 12 h) en las instalaciones de la casa de animales del instituto. Los animales fueron alimentados con alimento granulado estándar (Nutrimix Std-1020) obtenido de Nutrivet Life Sciences, Pune, India y se proporcionó agua a voluntad.Excluyendo el periodo de evaluación de parámetros de comportamiento. El estudio fue aprobado por el Comité Institucional de Ética Animal (Aprobación No. IAEC / RCPIPER / 2016-17 / 02) del Instituto RC Patel de Educación e Investigación Farmacéutica, Shirpur, India (Reg. No. 651 / PO / ReBi / S / 02 / CPCSEA). Todos los procedimientos experimentales que involucran el uso de animales se llevaron a cabo de acuerdo con las regulaciones establecidas por el Comité para el propósito del control y supervisión de la experimentación en animales (CPCSEA) constituida bajo la Ley de Prevención de la Crueldad a los Animales, 1960, Ministerio de Medio Ambiente. y Bosques, Gobierno de la India.
La inducción de la constricción crónica i njury (CCI) en ratas
La cirugía se realizó para inducir el ICC como se describió anteriormente por Chanchal et al . 5 . Brevemente, los animales fueron anestesiados con administración intraperitoneal de pentobarbital sódico (60 mg / kg). Se realizó una disección roma a través del bíceps femoral para exponer el nervio ciático común de la extremidad trasera derecha en la mitad del muslo. Aproximadamente, 5–7 mm del nervio se liberaron del tejido adherido proximal a la trifurcación del nervio ciático y cuatro ligaduras (6.0 seda) se ataron alrededor de 1 mm aproximadamente. Después de la ligadura nerviosa, las capas musculares y de la piel se suturaron instantáneamente y se aplicó externamente una solución de povidona y yodo. Las ratas se mantuvieron en jaulas individuales y se dejaron recuperar 29. Los tratamientos farmacológicos respectivos se iniciaron al día siguiente después de la cirugía.
Tratamiento de drogas y grupos.
Los animales se dividieron al azar en cinco grupos, cada uno de los cuales consistía en 8 ratas (n = 8). Grupo I: el grupo control normal de ratas se administró por vía oral una vez al día con 1 ml de solución salina durante 14 días. Grupo II: el grupo de ratas de operación simulada se trató con 1 ml de solución salina una vez al día durante 14 días. Grupo III: grupo de ratas con control de neuropatía inducida por CCI que recibió por vía oral 1 ml de solución salina una vez al día durante 14 días. Grupo IV: grupo de ratas tratadas con RT + de la neuropatía inducida por CCI que recibieron por vía oral 0,1 ml de RT ( dilución 1 × 10-12 ) con 1 ml de agua destilada una vez al día durante 14 días. Grupo V: grupo de ratas tratadas con neuropatía inducida por CCI + gabapentina que recibió por vía oral gabapentina (60 mg / kg / día, po) suspendida en carboximetilcelulosa (CMC) una vez al día durante 14 días.
Diseño experimental
Posterior a la inducción de ICC, las ratas se habituaron durante 3 días. El tratamiento con RT se inició al día siguiente después de la cirugía con ICC. La alodinia térmica y mecánica se midió el día 3, el día 7, el día 11 y el día 14 después de la cirugía mediante los métodos anteriores 5 , 40. La latencia de retirada de la pata (PWL) se registró con un tiempo de corte máximo de 20 segundos. La pata trasera derecha de cada rata hasta la articulación del tobillo se sumergió en agua tibia (40 ± 1 ° C) y agua fría (12 ± 1 ° C) para la determinación de la alodinia térmica (cálida y fría). La alodinia mecánica se observó utilizando un aparato electrónico Von-Frey que comprende sondas de punta (serie 2390, IITC Life Sciences Incorporation). El umbral de retirada de la pata (PWT) se registró con una presión de corte de 30 g. Las ratas se mantuvieron en jaulas de polipropileno con piso de malla metálica y se aclimataron durante aproximadamente 10 minutos antes de las mediciones. La superficie media plantar de la pata trasera operada se probó con filamentos de Von Frey a través del piso de malla, cuando la pata estaba en contacto con el piso. Cada filamento se aplicó a la superficie de la sembradora hasta que se dobló y se mantuvo en posición durante aproximadamente 6–8 segundos.55 .
El dia 14El día después de la cirugía, los animales se anestesiaron con inyección intraperitoneal de pentobarbital sódico (60 mg / kg). La temperatura corporal del animal se mantuvo a 37 ° C. La velocidad de conducción del nervio motor ciático-tibial (MNCV) se midió mediante la estimulación de los nervios ciático y tibial en la muesca ciática y la muesca tibial a través de los electrodos de aguja bipolar (Power Lab / ML856; AD Instruments, Australia) a la frecuencia de 0,10 Hz, 0,1 Duración de ms y amplitud de 1,5 V. Después de un estímulo único, el potencial de acción muscular compuesto se midió a partir del primer músculo interóseo de la pata trasera mediante electrodos de clavija unipolares. La grabación fue una respuesta bifásica típica con una onda M inicial que es una respuesta motora directa debido a la estimulación de las fibras motoras.32 , 56 .
Tras el registro de MNCV, las ratas se sacrificaron utilizando una sobredosis de pentobarbital sódico. El nervio ciático lesionado se aisló junto con segmentos de 1 cm en ambos lados de la lesión por ICC. La porción central de 5 mm del segmento nervioso aislado se procesó para examen histológico. Las secciones de 4 µm de espesor se tiñeron con hematoxilina y eosina. Las secciones teñidas se examinaron bajo el microscopio de luz en busca de alteraciones estructurales que incluyen desarreglo de la fibra, hinchamiento de la fibra nerviosa y presencia de células satélite activadas y células de Schwann. Los tejidos nerviosos fijados con paraformaldehído se deshidrataron en series graduales ascendentes de alcohol y se incluyeron en parafina. Las muestras se cortaron en secciones de 4 µm de grosor usando microtomo y se tiñeron con hematoxilina y eosina de acuerdo con los protocolos de tinción de rutina.13 , 57 .
Los segmentos de nervio ciático de las ratas se procesaron en tampón de fosfato enfriado con hielo (pH 7,4) para obtener el 10% de homogeneizado. El homogeneizado se centrifugó a 2000 g durante 20 min a 4 ° C y se utilizaron partes alícuotas para la determinación de malondialdehído (MDA) 58 , glutatión reducido (GSH) 59 , superóxido dismutasa (SOD) 38 y catalasa 39 , 60 .
El óxido nítrico (NO) se estimó utilizando el método descrito anteriormente por Kumar et al . 41 con algunas modificaciones. En resumen, se mezclaron 50 μl de tejido sobrenadante con 500 μl de reactivo de Griess y se determinó la absorbancia espectrofotométricamente a 540 nm utilizando un espectrofotómetro de microplaca Powerwave XS (Biotek, EE. UU.). La curva de calibración se obtuvo utilizando nitrito de sodio como estándar. La concentración de NO se expresó en μM de NO por mg de proteína.
Información Adicional
Nota del editor: Springer Nature se mantiene neutral respecto a las reclamaciones jurisdiccionales en mapas publicados y afiliaciones institucionales.
Cambia la historia
01 de octubre de 2018
Nota de los editores: se advierte a los lectores que las conclusiones de este documento están sujetas a críticas que están siendo consideradas por los editores. Se tomarán las medidas editoriales apropiadas una vez que se resuelva este asunto.
Referencias
- 1.Colloca, L. et al . Dolor neuropático. Nature Reviews Disease Primers 3 , 17002 (2017).
- 2.Liu, N. y col . Efectos antinociceptivos del gentiopicrósido en el dolor neuropático inducido por la lesión crónica por constricción en ratones: un estudio conductual y electrofisiológico. Revista canadiense de fisiología y farmacología 94 , 769–778 (2016).
- 3.Kim, HJ Berberine mejora la alodinia inducida por la lesión crónica por constricción del nervio ciático en ratas. Revista de alimentos medicinales 18 , 909–915 (2015).
- 4.Shahid, M., Subhan, F., Ahmad, N. y Ullah, I. Un extracto de Bacosides que contiene Bacopa monnieri alivia la alodinia y la hiperalgesia en el modelo de lesión por constricción crónica del dolor neuropático en ratas. BMC medicina complementaria y alternativa 17 , 293 (2017).
- 5.Chanchal, SK et al . Efectos protectores in vivo e in vitro del omeprazol contra el dolor neuropático. Informes científicos 6 , 30007 (2016).
- 6.Attal, N. Tratamiento farmacológico del dolor neuropático. Acta Neurologica Belgica 101 , 53–64 (2001).
- 7.Cohen, SP y Mao, J. Dolor neuropático: mecanismos y sus implicaciones clínicas. BMJ: British Medical Journal (en línea) 348(2014).
- 8.Jia, S., Zhang, Y. y Yu, J. Efectos antinociceptivos de la isosakuranetina en un modelo de rata de neuropatía periférica. Farmacología 100 , 201-207 (2017).
- 9.Kishore, L. y Singh, R. Efectos de diferentes potencias homeopáticas de Cephalendra indica en el tratamiento del dolor neuropático en la diabetes inducida por estreptozotocina. Boletín de la Facultad de Farmacia, Universidad de El Cairo 55 , 273–280 (2017).
- 10.Mohammadi, R., Amini, K. y Charehsaz, S. Tratamiento homeopático para la regeneración de nervios periféricos: un estudio experimental en un modelo de transección del nervio ciático de rata. Homeopatía 101 , 141–146 (2012).
- 11.Boyd, A. et al . Medicamentos o preparados a base de hierbas para el dolor neuropático. La Biblioteca Cochrane (2017).
- 12.Boyd, A. et al . Medicamentos o preparados a base de hierbas para el dolor neuropático y la fibromialgia. Base de Datos Cochrane de Revisiones Sistemáticas 5 (2013).
- 13.Muthuraman, A. y Singh, N. El efecto atenuante del extracto de Acorus calamus en la lesión por constricción crónica indujo dolor neuropático en ratas: una evidencia de efectos antioxidantes, antiinflamatorios, neuroprotectores e inhibidores del calcio. BMC medicina complementaria y alternativa 11 , 24 (2011).
- 14.Kaur, G., Bali, A., Singh, N. y Jaggi, AS Potencial de mejora de Ocimum sanctum en el dolor neuropático inducido por lesión por constricción crónica en ratas. Anais da Academia Brasileira de Ciências 87 , 417–429 (2015).
- 15.Zhou, J. et al . Paeoniflorin y albiflorin atenúan el dolor neuropático a través de la vía mapk en ratas con lesión por constricción crónica. Medicina complementaria y alternativa basada en evidencia 2016 (2016).
- dieciséis.Nurmikko, TJ et al . Sativex trata con éxito el dolor neuropático caracterizado por alodinia: un ensayo clínico aleatorizado, doble ciego, controlado con placebo. Pain® 133 , 210–220 (2007).
- 17.Babbar, S. et al . Análisis farmacocinético de la capsaicina después de la administración tópica de un parche de capsaicina de alta concentración a pacientes con dolor neuropático periférico. Control terapéutico de drogas 31 , 502–510 (2009).
- 18.Fisher, P. & Scott, D. Un ensayo controlado aleatorio de homeopatía en la artritis reumatoide. Rheumatology 40 , 1052-1055 (2001).
- 19.Dos Santos, A., Perazzo, F., Cardoso, L. y Carvalho, J. Estudio in vivo del efecto antiinflamatorio de Rhus toxicodendron. Homeopatía 96 , 95-101 (2007).
- 20.Verma, P. & Vaid, I. (Jain Publishers, Nueva Delhi, India, 2002).
- 21.Patil, C. et al . Actividad inmunomoduladora de Toxicodendron pubescens en modelos experimentales. Homeopathy 98 , 154-159 (2009).
- 22.Patil, CR et al . Efecto doble de Toxicodendron pubescens sobre el edema de la pata inducido por carragenina en ratas. Homeopathy98 , 88–91 (2009).
- 23.Patel, DR et al . Toxicodendron pubescens conserva su eficacia antiartrítica en diluciones homeopáticas 1M, 10M y CM. Homeopathy 101 , 165–170 (2012).
- 24.Patil, CR et al . Modulación de la artritis en ratas por Toxicodendron pubescens y sus diluciones homeopáticas. Homeopatía 100 , 131-137 (2011).
- 25.Guimarães, FS et al . Estimulación de la actividad de los linfocitos anti-melanoma por macrófagos co-cultivados activados por medicamentos homeopáticos complejos. BMC cancer 9 , 293 (2009).
- 26.Karp, J.-C. et al . El tratamiento con Ruta graveolens 5CH y Rhus toxicodendron 9CH puede reducir el dolor y la rigidez en las articulaciones relacionados con los inhibidores de la aromatasa en mujeres con cáncer de mama precoz: resultados de un estudio observacional piloto. Homeopatía 105 , 299–308 (2016).
- 27.Chen, L., Chen, W., Qian, X., Fang, Y. y Zhu, N. Liquiritigenin alivia la hiperalgesia mecánica y fría en un modelo de dolor neuropático de rata. Informes científicos 4 , 5676 (2014).
- 28.De Vry, J., Kuhl, E., Franken-Kunkel, P. y Eckel, G. Caracterización farmacológica del modelo de lesión por constricción crónica del dolor neuropático. Revista europea de farmacología 491 , 137–148 (2004).
- 29.Bennett, GJ y Xie, Y.-K. Una mononeuropatía periférica en ratas que produce trastornos de la sensación de dolor como los que se ven en el hombre. Pain 33 , 87-107 (1988).
- 30.Espinosa ‐ Juárez, JV, Jaramillo ‐ Morales, OA & López ‐ Muñoz, FJ El haloperidol disminuye la hiperalgesia y la alodinia inducidas por una lesión de constricción crónica. Farmacología básica y clínica y toxicología (2017).
- 31.Dowdall, T., Robinson, I. & Meert, TF Comparación de cinco modelos diferentes de rata de lesión de nervio periférico. Farmacología Bioquímica y comportamiento 80 , 93-108 (2005).
- 32.Morani, AS & Bodhankar, SL Efecto neuroprotector del acetato de vitamina E en modelos de mononeuropatía en ratas. Neuroanatomía 7 , 33-37 (2008).
- 33.Asiedu, M., Ossipov, MH, Kaila, K. & Price, TJ acetazolamida y midazolam actúan sinérgicamente para inhibir el dolor neuropático. Pain 148 , 302–308 (2010).
- 34.Bhat, RA et al . Efecto del ácido ursólico en la atenuación del dolor neuropático inducido por la lesión por constricción crónica en ratas. Farmacología fundamental y clínica 30 , 517–528 (2016).
- 35.Patil, KR et al . Los triterpenoides pentacíclicos inhiben la activación mediada por IKKβ de la vía NF-κB: evidencias in silico e in vitro . PloS one 10 , e0125709 (2015).
- 36.Shanbhag, D. & Khandagale, A. Detección y estandarización de la arjuna Terminalia utilizada como medicamento en la homeopatía utilizando el método HPTLC. Int J Anal Bioanal Chem 1 , 57–60 (2011).
- 37.Eruslanov, E. y Kusmartsev, S. En Protocolos avanzados en estrés oxidativo II 57–72 (Springer, 2010).
- 38.Marklund, S. y Marklund, G. Participación del radical anión superóxido en la autooxidación del pirogalol y un ensayo conveniente para la superóxido dismutasa. The FEBS Journal 47 , 469-474 (1974).
- 39.Aebi, H. In Methods in enzymology vol. 105 121-126 (Elsevier, 1984).
- 40.Goecks, CS et al . Evaluación de los parámetros oxidativos en la médula espinal de rata después de la constricción crónica del nervio ciático. Investigación neuroquímica 37 , 1952–1958 (2012).
- 41.Kumar, A., Meena, S. y Pottabathini, R. Efecto de Ashwagandha (Withania somnifera) contra la alteración crónica inducida por lesiones de comportamiento y alteraciones bioquímicas: posible afectación del mecanismo del óxido nítrico. Revista Internacional de Nutrición, Farmacología, Enfermedades Neurológicas 4 , 131 (2014).
- 42.Kim, HK et al . Las especies reactivas de oxígeno (ROS) desempeñan un papel importante en un modelo de rata de dolor neuropático. Dolor 111 , 116-124 (2004).
- 43.Freire, MAM, Guimarães, JS, Gomes-Leal, W. y Pereira, A. Modulación del dolor por óxido nítrico en la médula espinal. Fronteras en neurociencia 3 , 24 (2009).
- 44.Naik, AK, Tandan, SK, Kumar, D. & Dudhgaonkar, SP El óxido nítrico y sus moduladores en el dolor neuropático inducido por lesión por constricción crónica en ratas. Revista europea de farmacología 530 , 59–69 (2006).
- 45.Kumar, A., Meena, S., Kalonia, H., Gupta, A. y Kumar, P. Efecto del óxido nítrico en el efecto protector de la melatonina contra la constricción crónica La lesión del nervio ciático indujo dolor neuropático en ratas. (2011).
- 46.Naik, AK et al . Papel del estrés oxidativo en la fisiopatología de la neuropatía periférica y la modulación por N-acetil-l-cisteína en ratas. European Journal of Pain 10 , 573-573 (2006).
- 47.Bansode, VJ et al . Efecto mejorador del piruvato de etilo en el dolor neuropático inducido por la lesión crónica por constricción del nervio ciático. Indian Journal of Pain 28 , 82 (2014).
- 48.Khalil, Z. & Khodr, B. Un papel para los radicales libres y el óxido nítrico en la recuperación retardada en ratas de edad con lesión crónica de los nervios de constricción. Radical Radical Libre y Medicina 31 , 430–439 (2001).
- 49.Nazıroğlu, M., Dikici, DM & Dursun,. El papel del estrés oxidativo y la señalización de Ca2 + en las vías moleculares del dolor neuropático en la diabetes: enfoque en los canales de PRT. Investigación neuroquímica 37 , 2065-2075 (2012).
- 50.Shubayev, VI & Myers, RR Upregulation e interacción de TNFα y gelatinasas A y B en la lesión dolorosa del nervio periférico. Investigación del cerebro 855 , 83–89 (2000).
- 51.Andrade, P. et al . El alivio del factor de necrosis tumoral α del dolor neuropático inducido experimentalmente se asocia con la modulación de la expresión del receptor de TNF. Revista de investigación en neurociencia 92 , 1490–1498 (2014).
- 52.Singh, AK, Kumar, S. y Vinayak, M. Desarrollo reciente en el efecto antihiperalgésico de los fitoquímicos: acciones antiinflamatorias y neuro-moduladoras. Investigación de la inflamación , 1–22 (2018).
- 53.Piccinelli, AC et al . El limoneno reduce la hiperalgesia inducida por la gp120 y las citoquinas por la modulación de la expresión de IL-1β y proteínas en la médula espinal de ratones. Ciencias de la vida 174 , 28–34 (2017).
- 54.Jadhav, HP et al . Estandarización de la tintura madre homeopática de Toxicodendron pubescens y correlación de sus marcadores flavonoides con la actividad biológica. Homeopatía 105 , 48–54 (2016).
- 55.Tsuda, M. et al . Alodinia mecánica causada por la inyección intraplantar del agonista del receptor P2X en ratas: afectación de la señalización del receptor P2X2 / 3 heteromérico en neuronas aferentes primarias insensibles a la capsaicina. Journal of Neuroscience 20 , RC90 – RC90 (2000).
- 56.Kandhare, AD, Raygude, KS, Ghosh, P., Ghule, AE y Bodhankar, SL Efecto neuroprotector de la naringina por la modulación de biomarcadores endógenos en la neuropatía diabética dolorosa inducida por estreptozotocina. Fitoterapia 83 , 650–659 (2012).
- 57.Sudoh, Y. et al . Evaluación neurológica e histopatológica después de amitriptilina intratecal de alto volumen. Anestesia regional y medicamentos para el dolor 29 , 434–440 (2004).
- 58.Ohkawa, H., Ohishi, N. y Yagi, K. Análisis de peróxidos lipídicos en tejidos animales por reacción del ácido tiobarbitúrico. Bioquímica analítica 95 , 351–358 (1979).
- 59.Moron, MS, Depierre, JW & Mannervik, B. Niveles de glutatión, glutatión reductasa y actividades de glutatión S-transferasa en pulmón y hígado de rata. Biochimica et Biophysica Acta (BBA) -General Subjects 582 , 67–78 (1979).
- 60.Muthuraman, A. & Singh, N. Efecto neuroprotector del extracto rico en saponina de Acorus calamus L. en un modelo de rata de lesión crónica por constricción (CCI) del dolor neuropático inducido por el nervio ciático. Diario de etnofarmacología 142 , 723–731 (2012).
Expresiones de gratitud
Los autores reconocen al laboratorio de farmacopea homeopática, Ghaziabad, India, por la generosa muestra de regalo de la muestra autenticada Toxicodendron pubescens . Los autores también reconocen el apoyo financiero a los trabajos de investigación en el laboratorio del Dr. Shreesh Ojha de la Universidad de los Emiratos Árabes Unidos, Emiratos Árabes Unidos.
Información del autor
Afiliaciones
Departamento de Farmacología, RC Patel Instituto de Educación e Investigación Farmacéutica, Shirpur-425405, Dist. Dhule, Maharashtra, India
- Shital Magar
- Umesh B. Mahajan
- Kalpesh R. Patil
- Sachin D. Shinde
- & Chandragouda R. Patil
Escuela de Biotecnología, Instituto de Tecnología Industrial de Kalinga (considerada universidad), Campus-11, Patia, Bhubaneswar, Odisha, Pin-751024, India
- Deepika Nayak
- Y Chanakya Nath Kundu
Instituto de Farmacia de SVKM, Dhule-424001, Dist-Dhule, Maharashtra, India
- Sameer N. Goyal
Centro de Homeopatía y Bienestar Janmangal, Bopal, Ahmedabad, Gujarat, 380058, India
- Shivang Swaminarayan
Departamento de Farmacología y Terapéutica, Facultad de Medicina y Ciencias de la Salud, Universidad de los EAU, Al Ain, EAU
- Shreesh ojha
Contribuciones
Concebió y diseñó los experimentos: CRP y CNK. Realizó los experimentos: SM, DN, UBM y SDS. Analizó los datos: SM, DN, SNG, CRP, UBM, CNK y SO. Reactivos, materiales y herramientas de análisis: CNK, CRP, SO y SNG escribió el documento: CNK, CRP, SM, KRP, SS y UBM escribió el primer borrador: SM, KRP, DN, CNK y UBM analizaron los datos, recopilaron referencias y redactaron el manuscrito: CRP, SO, CNK, SNG, KRP, SS y UBM
Conflicto de intereses
Los autores declaran no tener conflictos de intereses.
1 comentario:
The following time I learn a blog, I hope that it doesnt disappoint me as much as this one. I mean, I know it was my option to learn, however I truly thought youd have something fascinating to say. All I hear is a bunch of whining about one thing that you would fix in case you werent too busy on the lookout for attention. online casino games
Publicar un comentario